
 French
French Deutsch
DeutschParingstype

1. Knopvorming
2. Versmelting
3. Spore
Een paringstype is bij schimmels en waterschimmels een moleculair incompatibiliteitssysteem, dat onafhankelijk van de sekse paring van dezelfde paringstypen verhindert. De verschillende paringstypen worden afhankelijk van de groep weergegeven met cijfers, letters of met "+" en "–" in plaats van mannelijk (♂) en vrouwelijk (♀).
Als er van geslachten/seksen gesproken wordt, dan wordt de ouder die de celkern geeft, aangeduid als "mannelijke" en de ouder die de celkern ontvangt als de "vrouwelijke". Schimmels die alleen celkernen geven of die alleen celkernen ontvangen worden net zoals bij planten tweehuizig genoemd. Geven ze celkernen en ontvangen ze ook celkernen dan heten ze eenhuizig.
Of er versmelting van de celkernen (karyogamie) plaatsvindt na de versmelting van de cellen (plasmogamie), hangt af van de herkomst van de schimmeldraden (hyfen) of de gistcellen. Bij veel schimmels versmelten hyfen alleen als ze bij hetzelfde organisme horen. Er treedt dan zelfbevruchting op, een eigenschap die als 'homothallie' bekendstaat. Andere schimmels hebben een genetisch niet-identieke partner nodig en zijn daarmee 'heterothallisch'. Daarnaast moeten de twee betreffende hyfen ook nog passende paringstypen hebben. Deze zijn een bijzonderheid van schimmels en waterschimmels en kunnen vergeleken worden met geslachten bij planten en dieren, met dat verschil dat er bij schimmels meer dan twee paringstypen kunnen zijn. Bij de basidiomyceten komen meer dan duizend paringstypen voor.[1]
Ascomyceten
[bewerken | brontekst bewerken]Genetische achtergrond
[bewerken | brontekst bewerken]
Genen voor paringstype bij de ascomyceten worden wel idiomorfen in plaats van allelen genoemd vanwege de onzekerheid over de gemeenschappelijke voorouder. De door de genen gecodeerde proteïnen zijn transcriptiefactoren, die de vroege en de late stadia ven de seksuele cyclus reguleren. Heterothallische ascomycetes produceren gameten met een enkel Mat-idiomorf (allel) en versmelting is alleen mogelijk tussen gameten met complementaire paringstypen.
Ascomyceten hebben gewoonlijk twee paringstypen: "MAT1-1" en "MAT1-2", in vergelijking met die van de gistlocus MAT.[2] MAT1-1 (MAT1) codeert voor het regulatoreiwit met een DNA-bindingsmotief, terwijl MAT1-2 (MAT2) codeert voor een proteïne met een alfaboxmotief, zoals in het gist paringstype MATα1.[3] De overeenkomstige paringstypen bij gist worden aangeduid met MATa en MATα. Het DNA-bindingsmotief is elektroforetisch zeer beweeglijk en wordt daarom aangeduid met HMG (High Mobility Group).
Homothallische ascomyceten produceren gameten, die met elke andere gameet in de populatie kan versmelten inclusief de eigen mitotisch gevormde gameten doordat vaak elke haploïde twee verschillende vormen van de Mat-locus in hun genoom hebben.[4]
De genen die het paringstype bepalen liggen in de homeobox en coderen voor enzymen, die feromonen en feromoonreceptoren produceren. De geslachtelijke voortplanting hangt af van de door verschillende allelen van hetzelfde gen geproduceerde feromonen. Daar deze gebeurt bij haploïden, zijn door versmelting van cellen of hyfen, complementaire genen van een geschikte partner noodzakelijk. Het aantal paringstypen is afhankelijk van het aantal genen en het aantal allelen per gen. Afhankelijk van de schimmelsoort vindt geslachtelijke voortplanting plaats door gameten dan wel versmelting van hyfen. Wanneer een receptor van een haploïde een feromoon van een complementair paringstype waarneemt, vindt chemotropische groei of als het een gameet is chemotactische beweging plaats.[5]
Paring
[bewerken | brontekst bewerken]Bij compatibiliteit vormen zich op de schimmeldraden gametangiën, de bouwcellen voor de gameten, waarin zich talrijke celkernen verzamelen. Uit een zeer fijne, vertakte, gespiraliseerde, meercellige schimmeldraad, de trichogyne, die uit een van de gametangiën, nu ascogonium (vrouwelijk) genoemd, ontstaat, vormt zich een verbinding waardoor de celkernen uit het andere gametangium, nu antheridium (mannelijk) genoemd, zich naar het ascogonium kunnen bewegen.
De opeenvolgende stappen in het fusieproces van A en a haploide cellen zijn beschreven door Fincham and Day[6] en Wagner en Mitchell.[7] Na de fusie van de cellen volgt niet direct de fusie van de kernen. Eerst gaan de bij elkaar horende kernen van de bevruchtende cel (het antheridium) en die van het ascogonium zich synchroon delen. Vervolgens kromt de top en vormt zo aan de top een haak in de vorm van een bisschopsstaf. Daarna worden er twee tussenwanden gevormd. De middelste cel bevat één kern type A en één kern type a. Deze tweekernige cel start de vorming van een ascus en wordt de “initiële ascuscel” genoemd. De twee overgebleven eenkernige cellen smelten vervolgens samen. Deze tweekernige cel kan op zich ook weer haakvormig worden en een eigen initiële ascuscel vormen. Dit proces kan zich vele malen herhalen.
Na de vorming van de initiële ascuscel smelten de twee kernen samen en vormen een diploïde cel. Dit is het enige diploïde stadium van de schimmel. De diploïde kern bevat 14 chromosomen. Direct na de vorming van de diploïde kern gaat deze zich delen door een eerste meiotische deling en worden er na een tweede meiotische deling vier haploïde kernen gevormd, twee van het type A en twee van het type a. Na vervolgens een mitotische deling zijn er in elke ascus vier kernen van het type A en vier kernen van het type a.
-
 Levenscyclus van Neurospora crassa
Levenscyclus van Neurospora crassa

Basidiomyceten
[bewerken | brontekst bewerken]
a= aecium; sp= spermogonium;
ur= uredospore; t= teleutospore
In het algemeen komt zowel 'heterothallie' als 'homothallie' voor bij de basidiomyceten. Heterothallie komt bij de basidiomyceten het meeste voor. Ongeveer 90% van de Agaricomycotina zijn heterothallisch.[8]
Sommige soorten hebben van alle schimmels de meest complexe geslachtelijke voortplanting. Zo heeft zwarte roest (Puccinia graminis) een monocotyle waardplant voor de vegetatieve fase en een dicotyle waardplant voor de generatieve fase. De schimmel vormt vijf verschillende sporen: uredosporen, teleutosporen, basidiosporen, spermatiën en aecidiosporen.
De paringscompatibiliteit wordt bij de basidiomyceten in twee paringstypen ingedeeld: tetrapolair en bipolair.
- Een bipolaire schimmel heeft één gen voor paring met de allelen A of a.
- Een tetrapolaire schimmel heeft twee, onafhankelijke genen voor paring. Ze worden met A en B (bij de onderstam Agaricomycotina) of a en b (bij de onderstammen Ustilaginomycotina en Pucciniomycotina) aangeduid. Het a gen heeft twee allelen en het b gen meer dan twee. De twee basidiosporen (sporidia) moeten verschillende allelen van beide genen hebben willen ze zich met elkaar verenigen. Zo is een basidiospore met a1b2 verenigbaar met een basidiospore met a2b1 allelen. Het a gen codeert voor een feromoon, waardoor de vorming van een conjugatiebuis en versmelting in gang gezet wordt. Het b gen codeert voor regulerende eiwitten die zich aan het DNA-binden. Er kunnen meer dan twee allelen per gen voorkomen. Builenbrand heeft meer dan 25 b, maar slechts 2 a allelen.[9] Coprinopsis cinerea heeft meer dan 240 allelen van zowel A als B en Schizophyllum commune heeft meer dan 339 allelen van A en 64 van B.[10]
Alleen schimmels met verschillende allelen zijn compatibel en kunnen paren. De paring begint met de vorming van een conjugatiebuis en de uitwisseling van celkernen. Er ontstaan dikaryotische (tweekernige) hyfen met de afzonderlijke haploïde kernen van beide ouders. Onder de juiste milieuomstandigheden wordt er een vruchtlichaam met basidia op een hymenofoor gevormd. De hymenofoor is de drager van de basidia op het hymenium. Een basidium ontstaat meestal door gespvorming. Daarnaast zijn er ook soorten die geen gespvorming hebben. Hier ontstaat een basidium door knopvorming van de eindcel aan een dikaryotische hyfe. Een derde variant is de vorming van een probasidium.
Op het basidium vindt de vorming van basidiosporen (sporidia) plaats via karyogamie en meiose. De dikaryotische hyfen behouden vaak de gesp-verbinding. De vorming van deze verbindingen wordt gereguleerd door de beide paringsgenen. Voorbeelden zijn de branden, zoals builenbrand (Ustilago maydis).[11][12] en de paddenstoel waaiertje (Schizophyllum commune).[13]
-
 Paring van twee compatibele monokaryons bij de Basidiomycota
Paring van twee compatibele monokaryons bij de Basidiomycota -
 gespvorming
gespvorming -




De generatieve fase van de roesten vindt vaak plaats op een tussenwaardplant. Bij zwarte roest is dit de zuurbes (Berberis vulgaris). Hier vindt de generatieve fase plaats, waarbij spermatiën en aecidiosporen gevormd worden. De teleutospore is de dikwandige, gekleurde rustspore, die in de loop van het seizoen gevormd wordt in het telium (vruchtlichaam) op de andere waardplant. Het telium wordt net voor het afsterven van deze waardplant gevormd. De gesteelde teleutospore is tweecellig en heterothallisch. In het voorjaar kiemt de teleutospore en vormt een basidium, waarop de basidiosporen gevormd worden. De basidiospore kiemt op berberis en vormt daar een spermogonium met spermatiën en receptieve hyfen. Na bevruchting van een receptieve hyfe van het spermogonium door een spermatium met een ander paringstype wordt een aecium met haploïde-dikaryotische aecidiosporen gevormd. Ook kan er een aecium gevormd worden uit twee paringshyfen met verschillend paringstype. De aecidiospore kiemt op de graanplant, waarna een uredinium gevormd wordt. Het uredinium vormt uredosporen, die vervolgens weer een uredinium vormen. Later in het seizoen gaat een uredinium over in een telium en gaat teleutosporen produceren of vormt het mycelium een telium.
-
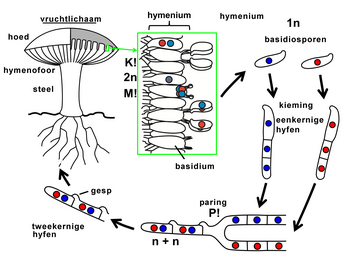 Levenscyclus Agaricus-soorten, zoals van de champignon. P=paring, K=karyogamie, M=meiose.
Levenscyclus Agaricus-soorten, zoals van de champignon. P=paring, K=karyogamie, M=meiose. -
 Levenscyclus builenbrand
Levenscyclus builenbrand -
 Levenscyclus zwarte roest
Levenscyclus zwarte roest
_nl_txt.png)
.jpg)

Aantasting van sperzieboon en pronkboon door roest (Uromyces appendiculatus var. appendiculatus).
- Uromyces appendiculatus var. appendiculatus
-
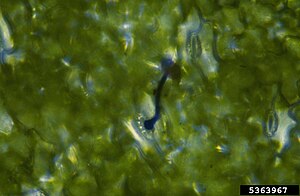 Kiembuis uredospore met appressorium
Kiembuis uredospore met appressorium -
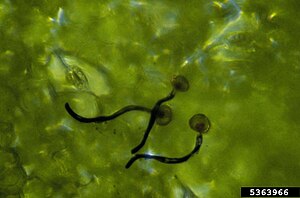 Kiembuizen uredosporen
Kiembuizen uredosporen -
 Uredosporen (licht gekleurd) en teleutosporen (donkergekleurd)
Uredosporen (licht gekleurd) en teleutosporen (donkergekleurd) -
 Telia
Telia -
 Telia
Telia -
 Telia
Telia -
 Telia
Telia -
 Spermogonia (links) en Aecidia (rechts)
Spermogonia (links) en Aecidia (rechts) -
 Spermogonia
Spermogonia -
 Spermogonium
Spermogonium -
 Aecidia op de peul van pronkboon
Aecidia op de peul van pronkboon










.jpg)
Oömyceten
[bewerken | brontekst bewerken]De waterschimmels, Oomycota (Winter 1879), behoren niet tot de schimmels, maar tot het rijk Chromista, en zijn meer verwant aan de diatomeeën en de zonnediertjes.
Bij de waterschimmels worden na bevruchting van een oögonium van bijvoorbeeld het type A1 met een antheridium van bijvoorbeeld het type A2 oösporen gevormd.
Levenscyclus aardappelziekte
[bewerken | brontekst bewerken]De veroorzaker van de aardappelziekte, Phytophthora infestans overwintert gewoonlijk in besmette knollen die achterblijven op het land. Uit de geïnfecteerde knollen ontwikkelen zich planten die de ongeslachtelijke sporen (sporangia en zoösporen) opleveren. Uit deze sporen kunnen nieuwe infecties ontstaan als het gewas ten minste gedurende vier à acht uur nat blijft, de zogenaamde bladnat-periode, bij een relatieve luchtvochtigheid van meer dan 95%. Na infectie ontstaan bij een temperatuur van 12-24 graden Celsius binnen enkele dagen met het oog waarneembare symptomen op bladeren en stengels. Na binnendringing van de oömyceet in de plant duurt het 3 tot 5 dagen voordat er nieuwe sporen gevormd worden.
Tot de jaren tachtig was in Europa alleen het zogenaamde A1 type aanwezig. Door de introductie van nieuwe A1 en A2 paringstypen in Europa is nu ook de geslachtelijke fase met oösporen aanwezig, die in tegenstelling tot de sporangia en zoösporen gedurende een lange periode buiten de waardplant in leven blijven en minstens drie jaar in de grond overleven. De zoösporen spelen waarschijnlijk pas later in het seizoen (eind juni - begin juli) een rol. Bij de paring van een hyfe van het type A1 met een hyfe van het type A2 worden een oögonium en een antheridium gevormd, waaruit de oöspore ontstaat. Deze eicel wordt binnen een gametangium door een spermatozoïde bevrucht. Uit de oösporen ontstaan sporangia en zoösporangia. De zoösporangia vormen ten slotte de zoösporen.

-
 Phytophthora voortplantingsorganen: A: sporangium. B: zoöspore. C: chlamydospore. D: oöspore.
Phytophthora voortplantingsorganen: A: sporangium. B: zoöspore. C: chlamydospore. D: oöspore. -
 Sporangium en zoöspores
Sporangium en zoöspores


Zygomyceten
[bewerken | brontekst bewerken]Paringshyfen, bij de zygomyceten zygoforen genoemd, met twee verschillende paringstypen, (+) en (-), groeien onder invloed van de gevormde, vluchtige feromonen door chemotropie naar elkaar toe. De feromonen bestaan hoofdzakelijk uit trisporisch zuur en de precursors. Trisporisch zuur is een C-18 terpenoïde, dat gevormd wordt via β-caroteen en retinol reactiepaden.[14] Bij contact plakken de celwanden aan elkaar, vlakken af en ontstaat er een fusiescheidingswand. De toppen van de zygofoor zwellen knotsvormig op en vormen uiteindelijk met de progametangia een brug. Bij de toppen van de twee knotsvormige cellen wordt een scheidingswand gevormd, die het terminale gametangium afscheiden van het progametangium. In dit stadium worden de buitenste cellen een suspensor genoemd, die voor ondersteuning zorgen. De middelste cellen gaan de zygospore vormen. Blaasjes verzamelen zich bij de tussenwand van de twee middelste cellen, die vervolgens begint op te lossen en de protoplasma's gaan zich vermengen. Er vindt vervolgens plasmogamie plaats. Vlak voordat de tussenwand geheel is opgelost, gaan de buitenste celwanden zich verdikken. Dit is te zien als donkere vlekken op de buitenste celwanden. Deze donkere vlekken kunnen uitgroeien tot wratachtige structuren, die de dikte van de wand van de zygospore bepalen. Tijdens de groei van de zygospore worden de wratachtige structuren ook groter en zitten dan uiteindelijk rondom de gehele cel. Nu kunnen de stralen van een elektronenmicroscoop niet langer door de wand dringen. Er zijn soorten die naakte zygosporen en soorten die ingekapselde zygosporen vormen.
Meiose vindt gewoonlijk plaats voordat de zygospore gaat kiemen. Het tijdstip van meiose kan in drie hoofdtypen onderscheiden worden.
- Type 1. De kernen versmelten snel, binnen enkele dagen.
- Type 2. Sommige kernen degenereren en de meiose vindt pas plaats bij de kieming.
- Type 3. De haploïde kernen splitsen zich continu door mitose. Vervolgens vormen sommige kernen groepen. Er zijn zowel diploïde als haploïde kernen in het sporangium aanwezig.
-
 Zygosporevorming bij Rhizopus nigricans
Zygosporevorming bij Rhizopus nigricans -
 Naakte zygospore (meiospore: door meiose ontstane spore)
Naakte zygospore (meiospore: door meiose ontstane spore) -
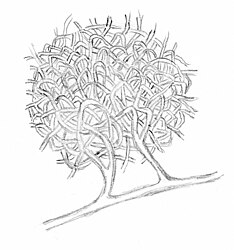 Ingekapselde zygospore
Ingekapselde zygospore -
 Plasmogamie bij de vorming van een Rhizopus-zygospore van een broodschimmel
Plasmogamie bij de vorming van een Rhizopus-zygospore van een broodschimmel -
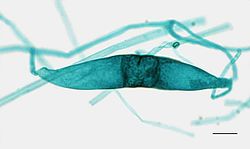 Later stadium van plasmogamie bij de vorming van een Rhizopus-zygospore van een broodschimmel
Later stadium van plasmogamie bij de vorming van een Rhizopus-zygospore van een broodschimmel -
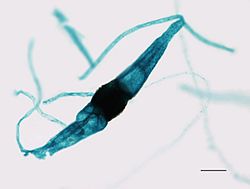 Onvolgroeide Rhizopus-zygospore van een broodschimmel
Onvolgroeide Rhizopus-zygospore van een broodschimmel -
 Volgroeide Rhizopus-zygospore van een broodschimmel
Volgroeide Rhizopus-zygospore van een broodschimmel -
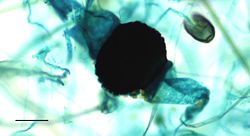 Kiemende Rhizopus-zygospore van een broodschimmel
Kiemende Rhizopus-zygospore van een broodschimmel









- ↑ Casselton LA, Mate recognition in fungi, Heredity, 2002, 88, 2, 142–147, PMID 11932772, DOI:10.1038/sj.hdy.6800035
- ↑ Yoder, O.C., Valent, Barbara, Chumley, Forrest, Genetic Nomenclature and Practice for Plant Pathogenic Fungi, Phytopathology, 1986, 76, 4, 383–385. [1], geraadpleegd 11 november]
- ↑ Turgeon, B.G., Yoder, O.C., Proposed Nomenclature for Mating Type Genes of Filamentous Ascomycetes, Fungal Genetics and Biology, 2000, 31, 1–5, DOI:10.1006/fgbi.2000.1227
- ↑ Giraud, T., Mating system of the anther smut fungus Microbotryum violaceum: Selfing under heterothallism, Eukaryotic Cell, 2008, 7, 765–775, DOI:10.1128/ec.00440-07
- ↑ Polaino, S., Idnurm, A. (2012). Sexual Pheromones in the Fungi. In: Witzany, G. (ed). Biocommunication of Fungi. Springer Science + Business Media. pp 171-188. ISBN 978-94-007-4263-5.
- ↑ Fincham J RS, Day PR (1963). Fungal Genetics. Blackwell Scientific Publications, Oxford, UK. ASIN: B000W851KO
- ↑ Wagner RP, Mitchell HK. (1964). Genetics and Metabolism. John Wiley and Sons, Inc., New York ASIN: B00BXTC5BO
- ↑ James, Timothy, Sex in Fungi: Molecular Determination and Evolutionary Implications, 2007, ASM Press, Washington DC, 317–331, J. Heitman, J. W. Kronstad, J. W. Taylor and L. A. Casselton, Analysis of mating type locus organization and synteny in mushroom fungi: Beyond model species
- ↑ Hsueh, Y. P., Heitman, J., Orchestration of sexual reproduction and virulence by the fungal mating-type locus, Current Opinion in Microbiology, 2008, 11, 6, 517–524, DOI:10.1016/j.mib.2008.09.014
- ↑ Casselton, L. A., Kües, U., J. Heitman, Sex in Fungi: Molecular Determination and Evolutionary Implications, 2007, ASM Press, Washington, DC, 283–300, editors: J. W. Kronstad, J. W. Taylor, L. A. Casselton, The Origin of Multiple Mating Types in the Model Mushrooms Coprinopsis cinerea and Schizophyllum commune
- ↑ Bakkeren, G, Kronstad, J. W., The Pheromone Cell Signaling Components of the Ustilago a Mating-Type Loci Determine Intercompatibility Between Species, Genetics, 1996, 143, 1601–1613
- ↑ Schulz, B, Banuett, F Dahl, M Schlesinger, R Schäfer, W Martin, T Herskowitz, I Kahmann, R, The b alleles of U. maydis, whose combinations program pathogenic development, code for polypeptides containing a homeodomain-related motif, Cell, 1990, 60, 2, 295–306, DOI:10.1016/0092-8674(90)90744-y
- ↑ Raudaskoski, M, Kothe, E, Basidiomycete mating type genes and pheromone signaling, Eukaryotic, Cell, 2010, 9, 6, 847–859, DOI:10.1128/ec.00319-09
- ↑ Gooday, Graham W.; Carlile, Michael J., The discovery of fungal sex hormones: III. Trisporic acid and its precursors. Mycologist vol. 11 issue 3 August 1997. pp. 126–130
Literatuur
- Sitte, P., E. Weiler, J.W. Kadereit, A. Bresinsky, C. Körner: Lehrbuch der Botanik für Hochschulen. Begründet von Eduard Strasburger. 35. Auflage. Spektrum Akademischer Verlag, Heidelberg 2002, ISBN 3-8274-1010-X.
- Bakkeren, G. & J. W. Kronstad: Linkage of mating-type loci distinguishes bipolar from tetrapolar mating in basidiomycetous smut fungi In: Proceedings of the National Academy of Sciences USA. 91, Juli 1994, S. 7085-7089
- Fraser, J. A. & J. Heitman: Evolution of fungal sex chromosomes In: Molecular Microbiology. 51 (2), 2004, S. 299–306