
 French
French Deutsch
DeutschОогенез — Вікіпедія

Оогене́з або овогене́з (грец. ōón яйце + грец. genésis виникнення) — у тварин, розвиток жіночої статевої клітини — яйцеклітини (яйця).
Під час ембріонального розвитку організму гоноцити мігрують до зачатків жіночих статевих гонад (яєчників), де відбувається весь їхній подальший розвиток.
Оогенез поділяють на три етапи або періоди: розмноження, росту і дозрівання.
Потрапивши у яєчник, гоноцити стають оогоніями. Оогонії здійснюють період розмноження. У цей час оогонії діляться мітотичним шляхом.
Статеві клітини в цьому періоді називаються ооцитами першого порядку. Вони втрачають здатність до мітотичного поділу і входять у профазу I мейозу. У цей час відбувається ріст статевих клітин.
У періоді росту виділяють 2 стадії:
- Стадія малого росту (превітелоігенез) — об'єм ядра і цитоплазми збільшується пропорційно. При цьому ядерно-цитоплазматичне співвідношення не порушується. На цій стадії відбувається активний синтез всіх видів РНК — рибосомних, транспортних і матричних. Всі ці типи РНК синтезуються переважно про запас, тобто для використання вже заплідненою яйцеклітиною.
- Стадія великого росту (вітеліогенез) — об'єм цитоплазми ооциту може збільшитися в кілька десятків тисяч разів, тоді як об'єм ядра збільшується незначно. Отже, ядерно-цитоплазматичне співвідношення сильно зменшується. На цій стадії в ооциті I порядку утворюється жовток. За способом утворення розрізняють жовток екзогенний і ендогенний. Властивий більшості видів звірів екзогенний жовток утворюється на основі білка-попередника вітелогеніну, що надходить в ооцит ззовні. У хребетних вітелогенін синтезується в печінці матері та транспортується до фолікулу, що містить ооцит по кровоносним судинам. Потрапляючи потім у простір, безпосередньо навколо ооциту (периооцитний простір), вітелогенін поглинається ооцитом шляхом піноцитозу.
Дозрівання ооцита — це процес послідовного проходження двох поділів мейозу (поділів дозрівання). Як мовилося вище, в період до першого поділу мейозу ооцит тривалий час перебуває в стадії профази I мейозу, коли й відбувається його зростання. Вихід з профази I мейозу приурочений до досягнення самкою статевозрілості і визначається статевими гормонами.
Перший поділ у багатьох видів є редукційним, оскільки саме під час нього гомологічні хромосоми розходяться до різних клітин. Таким чином, кожна з клітин, що розділилися набуває половинного (гаплодїного) набору хромосом, де кожен ген представлений лише одним алелем. Кожна дочірня клітина після цього поділу має набір хромосом — n, і 2с — набір хроматид.
Оскільки першому діленню дозрівання передувала S-фаза, кожна з хромосом, які розійшлися, містить подвійну кількість ДНК, оскільки складається з двох хроматид. Ці генетично ідентичні хроматиди розходяться до різних сестринських клітин в другому поділі мейозу, що є екваційним (як і звичайний поділ соматичних клітин). Після двох поділів мейозу число хромосом у кожній з клітин є гаплоїдним (1n), а загальна кількість хроматид у кожному клітинному ядрі становитиме 1с.
Оогенез | | Дифузний <----|-----> Локалізований | | Солітарний <-----|-----> Аліментарний | | Нутриментарний <-----|-----> Фолікулярний
- Дифузний оогенез — розвиток яйцеклітини може відбуватись в будь-якій частині тіла (губки, кишковопорожнинні, війчасті черви). За дифузного оогенезу ооцити є фагоцитуючими клітинами, вони не синтезують і накопичують жовткові включення, а ростуть завдяки надходження низькомолекулярних сполук з фаголізосом. У цих ооцитах виробляється багато гідролітичних ферментів, що необхідні їм для фагоцитозу.
- Локалізований оогенез — розвиток яйцеклітини відбувається у жіночих гонадах — яєчниках.
- Солітарний оогенез — ооцит може розвиватися й без участі допоміжних клітин що постачають йому поживні речовини. Цей тип характерний для деяких кишковопорожнинних, червів, молюсків. У них ростучі статеві клітини позбавлені допоміжних елементів, жовточні білки і РНК синтезуються ними самостійно. Все необхідне для макромолекулярних синтезів ооцит отримує із зовнішнього середовища (порожнини гонади) у вигляді простих низькомолекулярних сполук.
- Аліментарний оогенез — розвиток ооциту відбувається за участі допоміжних харчуючих клітин.
- Нутриментарний оогенез — ооцит оточений трофоцитами (харчуючими клітинами), які зв'язані з ним цитоплазматическими містками (вищі черви, комахи). Трофоцити — це абортовані статеві клітини, тобто вони мають спільне походження з ооцитом, але їхній розвиток зупинився ще на етапі росту. На один ооцит припадає багато харчуючих клітин, що забезпечують статеву клітину РНК. У вітелогенезі трофоцити участі не беруть: жовток утворюється завдяки надходженню високомолекулярних речовин з ззовні.
- Фолікулярний оогенез — ооцит оточений фолікулярними клітинами соматичного походження, які утворюють з ним функціональну структуру — фолікул (переважна більшість тварин, зокрема всі хордові). Фолікулярні клітини не беруть участь у синтезі білків жовтка, всі види РНК синтезуються в самому ооциті. Виняток становлять фолікулярні клітини птахів і ящірок, які синтезують РНК для ооцита.
-
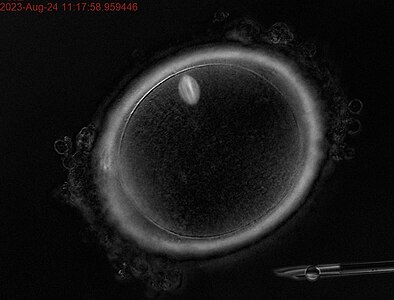 Ооцит людини на стадії метафази першого поділу мейозу. В позиції на 11 годину знаходиться веретено поділу.
Ооцит людини на стадії метафази першого поділу мейозу. В позиції на 11 годину знаходиться веретено поділу. -
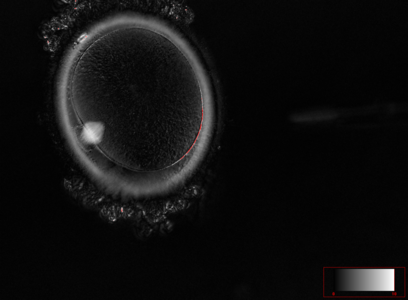 Ооцит людини на стадії анафази першого поділу мейозу. В позиції на 8 годину знаходиться веретено поділу (мейоз І), воно розміщене в місці відділення першого полярного тільця ооциту. Вловлено момент початку відділення першого полярного тільця.
Ооцит людини на стадії анафази першого поділу мейозу. В позиції на 8 годину знаходиться веретено поділу (мейоз І), воно розміщене в місці відділення першого полярного тільця ооциту. Вловлено момент початку відділення першого полярного тільця. -
 Ооцит людини на стадії телофази першого поділу мейозу. В позиції на 7 годину в перивітеліновому просторі знаходиться перше полярне тільце, але залишки веретена поділу ще зв’язують його з ооцитом, що вказує на фінальні етапи першого поділу мейозу.
Ооцит людини на стадії телофази першого поділу мейозу. В позиції на 7 годину в перивітеліновому просторі знаходиться перше полярне тільце, але залишки веретена поділу ще зв’язують його з ооцитом, що вказує на фінальні етапи першого поділу мейозу. -
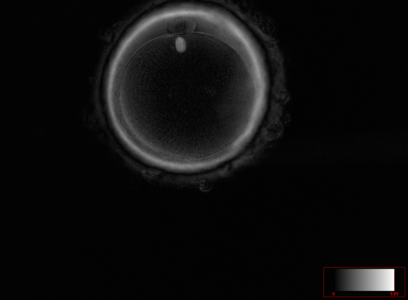 Ооцит людини на стадії метафази другого поділу мейозу. Зрілий та готовий до запліднення ооцит людини. В позиції на 12 годину в перивітеліновому просторі знаходиться перше полярне тільце. Під полярним тільцем в самому ооциті чітко візуалізується веретено другого поділу мейозу на стадії метафази. Візуалізується тришарова блискуча оболонка ооциту.
Ооцит людини на стадії метафази другого поділу мейозу. Зрілий та готовий до запліднення ооцит людини. В позиції на 12 годину в перивітеліновому просторі знаходиться перше полярне тільце. Під полярним тільцем в самому ооциті чітко візуалізується веретено другого поділу мейозу на стадії метафази. Візуалізується тришарова блискуча оболонка ооциту.




- ↑ X. Johne Liu (October 2012). Polar body emission. Cytoskeleton[en]. 69 (10): 670—685. doi:10.1002/cm.21041. PMID 22730245.
- Белоусов Л. В. Основы общей эмбриологии. — Москва: Издательство Московского университета : Наука, 2005. ISBN 5-211-04965-9(рос.)
- Голиченков В. А., Иванов Е. А., Никерясова Е. Н. Эмбриология. — Москва: Издательский центр «Академия», 2004. ISBN 5-7695-1168-0(рос.)
- Голиченков В. А., Иванов Е. А., Лучинская Н. Н. Практикум по эмбриологии. — Москва: Издательский центр «Академия», 2004. ISBN 5-7695-1330-6(рос.)
- Гилберт С. Биология развития. Т. 1-3 — Москва: «Мир» 1993—1995.(рос.)
| Довідкові видання | |
|---|---|
| Нормативний контроль |