
 French
French Deutsch
DeutschSilüriyen - Vikipedi
| Silüriyen | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
443,8 ± 1,5 - 419,2 ± 3,2 myö | |||||||||||||
 Erken Silüriyen'de Dünya (yaklaşık 440 myö) | |||||||||||||
| Kronoloji | |||||||||||||
| |||||||||||||
| Etimoloji | |||||||||||||
| Diğer telaffuz(lar) | Siluriyen | ||||||||||||
| Eş anlamlı(lar) | Gotlandiyen | ||||||||||||
| Kullanım bilgisi | |||||||||||||
| Gök cismi | Dünya | ||||||||||||
| Bölgesel kullanım | Küresel (ICS) | ||||||||||||
| Kullanılan zaman çizelgesi | ICS Zaman Cetveli | ||||||||||||
| Tanım | |||||||||||||
| Kronolojik birim | Dönem | ||||||||||||
| Stratigrafik birim | Sistem | ||||||||||||
| İlk öneren | Roderick Murchison, 1835 | ||||||||||||
| Zaman aralığının resmîyeti | Resmî | ||||||||||||
| Alt sınırını belirleyen | Bir graptolit olan Akidograptus ascensus'un ilk ortaya çıkışı. | ||||||||||||
| Alt sınır KSKN | Dob's Linn, Moffat, Birleşik Krallık 55°26′24″N 3°16′12″W / 55.4400°K 3.2700°B | ||||||||||||
| KSKN onayı | 1984[4][5] | ||||||||||||
| Üst sınırını belirleyen | Bir graptolit olan Monograptus uniformis'in ilk ortaya çıkışı | ||||||||||||
| Üst sınır KSKN | Klonk, Çekya 49°51′18″N 13°47′31″E / 49.8550°K 13.7920°D | ||||||||||||
| KSKN onayı | 1972[6] | ||||||||||||
| Atmosfer ve iklim verileri | |||||||||||||
| Deniz seviyesinin günümüze göre yüksekliği | Kısa dönemli negatif sapmalarla birlikte 180 metre civarında[7] | ||||||||||||
Silüriyen, Paleozoyik Zaman'ın en kısa süren jeolojik dönemi olarak bilinir. 443,8 milyon yıl önce başlayıp 419,2 milyon yıl önce sona ermiştir. Bu dönem, adını Galler'deki Silures kabilesinden alır. Silüriyen'in başlangıcı ve sonu için belirlenen kayaç tabakaları kesin olarak tanımlanmıştır ancak tam tarihler birkaç milyon yıl kadar hata payına sahiptir. Silüriyen Dönemi, Ordovisiyen-Silüriyen yok oluşu ile başlar; bu olay sırasında deniz canlılarının yaklaşık %60'ının nesli tükenmiştir.
Silüriyen, kara yaşamının ilk evrimleştiği dönem olarak yerküre tarihinde önem taşır. Damarlı bitkiler, ilkel karasal bitkilerden gelişmiş, dikarya mantarları çeşitlenmeye başlamıştır. Altı bacaklılar, örümceğimsiler ve çok bacaklılar tamamen karasal yaşama adapte olmuştur. Bu dönemde, çeneli ve kemikli balıkların evrimleşmesi de dikkate değer bir gelişmedir.
Silüriyen Dönemi'nin paleocoğrafyası, iklimi ve canlı türleri Paleozoyik Zaman'ın karmaşık ve dinamik yapısını yansıtır. Süperkıta Gondvana, ekvatorun üst yarısını ve Güney Yarımküre'nin büyük bir bölümünü kaplarken, Kuzey Yarımküre'nin çoğu büyük bir okyanusla kaplıydı. Yüksek deniz seviyeleri ve nispeten düz araziler, birçok ada topluluğunun ve çeşitli çevresel koşulların oluşmasına yol açtı. Silüriyen boyunca Gondvana yavaşça güneye doğru hareket etti, ancak bu dönemdeki buzulların Geç Ordovisiyen buzullarından daha az yayıldığına dair kanıtlar vardır. Yeni kıtasal kara kütleleri oluştuğunda deniz seviyesi yükseldi ve Silüriyen tortulları, erozyona uğramış Ordovisiyen tortullarının üzerine çökerek bir uyumsuzluk oluşturdu.
Silüriyen Dönemi, jeolojik ve biyolojik evrime dair kayda değer bilgiler sunar. İlk kara bitkilerinin ortaya çıkması, omurgalı balıkların çeşitlenmesi ve deniz canlılarının toprakta yaşamaya başlaması, bu dönemin kayda değer olaylarındandır. Dönemin kara ve deniz ekosistemleri, canlıların adaptasyonları ve çevresel değişimlerle etkileşim içinde şekillenmiştir. Silüriyen Dönemi'nin keşfedilmiş fosil kalıntıları, bu dönemin bilimsel anlayışına ışık tutarak tarih öncesi yaşamın evriminin anlaşılmasına yardımcı olmaktadır.
Kelime kökeni ve tarihi
[değiştir | kaynağı değiştir]Silüriyen sistemi, ilk kez 1830'ların başlarında Galler'in güneyindeki fosil taşıyan sedimanter kaya tabakalarını inceleyen bir İskoç jeolog olan Roderick Murchison tarafından tanımlandı. Bu katmanlara Galler'de yaşamış bir Kelt kabilesi olan Silures kabilesinin adını verdi. Murchison bu adlandırmayı, kendisinin üzerine çalıştığı jeolojik döneme Galler'in Latince ismi olan Cambria'dan esinlenerek "Kambriyen" adını koyan arkadaşı Adam Sedgwick'ten ilhâm alarak yapmıştır.[8] Ancak bu isimlendirmeye karşın, Silüriyen kayaçlarının oluşumu ve Silures kabilesinin eskiden üzerinde yaşamış olduğu topraklar arasında bir ilişki yoktur (kr Galler'in jeolojik haritası, Galler'in Roma İmparatorluğu öncesindeki haritası). 1835 yılında, bu iki jeolog "On the Silurian and Cambrian Systems, Exhibiting the Order in which the Older Sedimentary Strata Succeed each other in England and Wales" başlıklı ortak bir makale yayımlamıştır ve bu makale, modern jeolojik zaman cetvelinin temelini atmıştır.[9] İlk olarak tanımlandığı sıralar "Silüriyen" serisi daha uzak bölgelere kadar takip edildiğinde Sedgwick'in "Kambriyen" serisiyle çakışmaya başladı ve bu da arkadaşlığı sona erdiren anlaşmazlıklara neden oldu.
İngiliz jeolog Charles Lapworth, tartışmalı tabakaları içeren, Ordovisiyen adlı yeni bir sistem tanımlayarak çekişmeyi çözdü.[10] Silüriyen için alternatif adlandırmalardan biri de, Baltık adası Gotland'ın tabakalarından esinlenilmiş "Gotlandiyen" ismiydi.[11]
Fransız jeolog Joachim Barrande, Murchison'ın çalışmalarına dayanarak, Silüriyen terimini daha kapsamlı bir biçimde kullanmıştır ve kendinden sonra gelenler de Barrande'ın çalışmalarını haklı çıkarmıştır. Barrande, Bohemya'daki Silüriyen kayaçlarını sekiz kata böldü.[12] Bu yaptığı bölümleme 1854 yılında Edward Forbes tarafından sorgulandı[13] ve Barrande'ın Silüriyen'i ayırdığı sekiz kattan son üçü olan F, G ve H'nin, Devoniyen'e ait olması gerektiği bulundu.
Paleocoğrafya
[değiştir | kaynağı değiştir]
−4500 — – — – −4000 — – — – −3500 — – — – −3000 — – — – −2500 — – — – −2000 — – — – −1500 — – — – −1000 — – — – −500 — – — – 0 — |
| |||||||||||||||||||||||||||||||||||||||||||||
Süperkıta Gondvana, ekvatoru ve Güney Yarımküre'nin büyük bir bölümünü kaplarken, geniş bir okyanus da gezegenin kuzey yarısının çoğunu kaplıyordu.[14] Silüriyen Dönemi'nin yüksek deniz seviyeleri ve nispeten düz arazileri (kayda değer sıradağların az olduğu) bir dizi ada zinciri meydana getirdi ve bu nedenle zengin bir çevresel ortam çeşitliliği oluşturdu.[14]
İklim ve deniz seviyesi
[değiştir | kaynağı değiştir]
Silüriyen Dönemi'nde, bir zamanlar karasal buzulların hüküm sürdüğü Ordovisiyen ve Silüriyen'in ardından gelen Devoniyen'deki aşırı sıcaklıklara kıyasla nispeten istikrarlı ve ılık sayılabilecek sıcaklık seviyelerinin görüldüğü düşünülüyordu ancak şu anda, bu jeolojik dönem boyunca küresel iklimin birçok ciddi dalgalanma yaşadığı bilinmektedir.[15][16] Buna kanıt olarak bu dönemde birçok kayda değer karbon ve oksijen izotopu sapmaları bulunmaktadır.[17][18][19] Deniz seviyeleri, Silüriyen'in ilk yarısı boyunca Hirnasiyen'deki düşük seviyelerden yükseldi; ardından dönemin geri kalanı boyunca düştü.Daha küçük ölçekli modeller bu genel trendin üzerine bindirilmesine rağmen on beş kez yüksek deniz seviyesi (deniz seviyelerinin kıta sahanlığı kenarının üstünde olduğu dönemler) görülmüş olduğu belirlenmiştir ve Silüriyen'deki en yüksek deniz seviyesi muhtemelen bu dönem boyunca görülen en düşük seviyeye göre yaklaşık 140 metre (459 ft) daha yüksekti.[14]
Bu dönemde, Dünya sıcak bir sera evresine girdi. Bu sıcak sera evresinin oluşmasını sağlayan nedenler arasında atmosferde görülen 4500 ppm kadar yüksek CO2 seviyeleri ve ekvatordaki kara kütlerinin büyük bölümünü kaplayan sıcak ve sığ denizler vardır.[20] Silüriyen'in başlarında buzullar Güney Kutbu'na çekildi ve Silüriyen'in ortasına doğru neredeyse tamamen kayboldular.[16] Kırık kabuk tabakaları (coquina adlı), sıcak deniz yüzeylerinin tıpkı bugünkü gibi şiddetli fırtınalar ürettiği bir iklimin hâkim olduğuna dair güçlü kanıtlar sunar.[21]
Flora ve fauna
[değiştir | kaynağı değiştir]Silüriyen Dönemi, bazı paleontologlar tarafından Kambriyen ve Ordovisiyen'in büyük bölümü boyunca devam eden ardışık biyoçeşitlilik gelişiminin kesintiye uğradığı Geç Ordovisiyen kitlesel yok oluşunu (İngilizce LOME kısaltmasıyla anılır) takiben geniş bir iyileşme aralığı olarak görülmüştür.[22]
Silüriyen, göl, nehir kenarlarında ve büyük mikorizal nematofit ağlarında, Silüriyen-Devoniyen Karasal Devrimi'nin başlangıcını işaret eden yosun benzeri minyatür ormanlar şeklinde geniş karasal bitki örtüsü megafosillerinin görüldüğü ilk jeolojik dönemdir.[23][24][25] Ancak karada bulunan fauna, Devoniyen'de görülen çeşitlenmeden önce Dünya üzerinde kayda değer bir etki yaratmadı.[14]
Damarlı bitkilerin, yani su ve besin taşıyan dokulara sahip karasal bitkilerin ilk fosil kayıtları, Silüriyen Dönemi'nin ikinci yarısında görüldü.[26] Bu gruba ait en erken bilinen temsilciler Cooksonia'dır. Cooksonia'yı içeren çoğu tortul, denizel karakterdedir. Tercih ettikleri yaşam alanları muhtemelen nehir ve akarsu kenarlarıydı. Baragwanathia, Erken Ludlov'a (y. 420 milyon yıl öncesi) tarihlenen ve 10-20 santimetre (3.9-7.9 inç) uzunluğunda dallı saplar ve iğne benzeri yaprakları olan bir bitkidir. Bu bitki, fosil kalıntılarının yaşına göre yüksek bir gelişim derecesi gösterir. Bu bitkinin fosilleri Avustralya,[27][28] Kanada[29] ve Çin'de[30] keşfedilmiştir. Eohostimella heathana, Erken Silüriyen'deki (Landoveri) sıkışmış fosillerden bilinen,[31] muhtemelen karasal bir "bitkidir".[32] Bu bitkinin fosillerinin kimyası, algler yerine damarlı bitkilere benzer.[31]
Silüriyen Dönem'den karada yaşayan hayvan olarak kabul edilen fosiller de vardır. Bilinen en eski kırkayak, Geç Silüriyen'de (425 myö) Kerrera'da yaşamış Kampecaris obanensis ve Archidesmus türleridir.[33] Aynı zamanda Ludlov Devresi'nde (420 myö) yaşadığı bilinen kırkayaklar, çıyanlar ve trigonotarbid eklem bacaklılar da vardır.[33][34][35] Etçil omurgasızların varlığı, etçil olmayan av hayvanlarını da içeren basit besin ağlarının o zamanlarda da var olduğunu işaret etmektedir. 1990'da Andrew Jeram ve çalışma arkadaşları,[36] Erken Devoniyen biyotasından geriye doğru çıkarım yaparak mikroorganizmalarla beslenen, henüz keşfedilmemiş çökelciller ve otlayıcılar üzerine temellenen bir besin ağı ortaya koydu.[37] Cowie Formasyonu'nda keşfedilmiş Cowiedesmus ve Pneumodesmus gibi kırkayak türleri, 428 milyon yıl önce, yani Orta Silüriyen'deki en eski kırkayak türleri olarak kabul edilmişti.[38][39] Bununla birlikte, bu formasyonun yaşı bazı çalışmalar ile yeniden değerlendirilerek Erken Devoniyen'e tarihlendirildi.[40] Pneumodesmus, yine de hava ortamında solunum yapmak için stigma kullanan en eski kesin canlı kanıtı olması nedeniyle kayda değerdir.[33]
Osteichthyes adı verilen ilk kemikli balıklar Silüriyen'de ortaya çıktı. Kemikli pullarla kaplı dikenli yüzgeçliler bu balıklara örnektir. Balıklar dikkate değer ölçüde bir çeşitllendi ve önde bulunan iki ila üç solungaç yüzgecinin destek bölümüyle desteklenmiş hareketli çenelere sahip oldu. Birkaç metre uzunluğunda olan çeşitli öripterid türleri (deniz akrepleri), Kuzey Amerika'nın sığ Silüriyen denizleri ve göllerinde dolaşıyordu; bu canlıların fosillerinin birçoğu New York eyaletinde bulunmuştur. Sülükler de Silüriyen Dönemi'nde görülen canlılardandır. Brakiyopodlar bolca bulunuyordu ve çeşitlilerdi; Silüriyen brakiyopodları taksonomik bileşimi, ekolojisi ve çeşitliliğiyle Ordovisiyen'dekilerle benzerlik gösterir.[41] Geç Ordovisiyen kitlesel yok oluşundan sağ kurtulan brakiyopodlar, çevresel zorluklar için yeni adaptasyonlar geliştirdi[42] ve çoğu zaman soy tükenmesinin ardından tek bir paleolevhada endemik hale geldiler ancak daha sonra yayılım alanlarını genişlettiler.[43] En yaygın brakiyopodlar atripidler ve pentameridlerdi.[44] Atripidler Geç Ordovisiyen kitlesel yok oluşundan sonra Rudaniyen'de yeniden çeşitlenmeye ve çoğalmaya başlayan ilk gruptu,[45] pentamerid çeşitliliği ise ancak Aroniyen'de artmaya başladı.[44] Bryozoa, belirli bir sahil bölgesine dikkate değer oranda endemik hâle geldiler.[46] Ayrıca sölenterler[47] ve stromatolitlerle[48] simbiyotik ilişkiler geliştirdiler. Birçok çift çenetli fosili de Silüriyen yataklarında bulunmuştur[49] ve ilk su altında derinlere kazarak ulaşan çift çenetliler bu dönemdendir.[50] Kitonlar, Silüriyen'in ortasında biyoçeşitlilikte bir zirve yaşadı.[51] Hederelloidler, Silüriyen'de gelişti ve bazıları koloni yapan kıvrımlı mercan Entelophyllum ile simbiyozlar geliştirdi.[52] Silüriyen, tentakulitoidler için bir yükseliş dönemiydi; Baltoskandiya bölgesinde evrimsel bir patlama yaşadılar[53] ve Landoveri ile Venlok devrelerinde coğrafi aralıklarını genişlettiler.[54] Trilobitler, Rudaniyen'de toparlanmaya başladılar[55] ve Geç Ordovisiyen kitlesel yok oluşunun neden olduğu soy çeşitliliğindeki azalmaya rağmen, Ordovisiyen'deki gelişimlerini Silüriyen boyunca sürdürdüler.[56] Erken Silüriyen, Geç Ordovisiyen kitlesel yok oluşundan sonra yeniden çeşitlenen deniz zambakları için bir dönüşüm dönemiydi.[57] Geç Ordovisiyen kitlesel yok oluşundan en az etkilenen Flexibilia üyeleri, Silüriyen denizlerinde giderek artan bir ekolojik belirginlik kazandılar.[58] Monobatrid kamratlar, tıpkı flexibiller gibi, Landoveri'de çeşitlendiler ancak siyatokrinidler ve dendrokrinidler ise Silüriyen'in sonraki evrelerinde çeşitlendi.[59] Sayfokrinoid lobolitler Silüriyen-Devoniyen sınırından hemen önce ani bir şekilde ortaya çıktı ve bu ilk ortaya çıkışlarından çok kısa bir süre sonra Silüriyen sonlarında ortadan yok oldular.[60] Koralların ve stromatoporoidlerin endobiyotik simbiyontları yaygındı.[61][62] Özellikle kıvrımlı mercanlar, önceden belirtildiği gibi hederelloidler dahil olmak üzere[52] çeşitli epibiyontlarla kolonileşti ve kaplandı.[63]
Mercan resifleri aralıklı olarak yaygındı. Mercanlar, Silüriyen'de bazen fosillerde sık görülürken, bazen de kayaçlarda neredeyse yok denecek kadar az bulunmaktadır.[14]
Galeri
[değiştir | kaynağı değiştir]-
 Cooksonia, Orta Silüriyen'de görülen ilk damarlı bitkidir
Cooksonia, Orta Silüriyen'de görülen ilk damarlı bitkidir -
 Birleşik Krallık, Dudley, Wren's Nest Nature Reserve'den toplanmış Silüriyen'e tarihlenen deniz yatağı fosilleri
Birleşik Krallık, Dudley, Wren's Nest Nature Reserve'den toplanmış Silüriyen'e tarihlenen deniz yatağı fosilleri -

-
 Birleşik Krallık, Dudley, Wren's Nest Nature Reserve'den toplanmış Silüriyen'e tarihlenen deniz yatağı fosilleri
Birleşik Krallık, Dudley, Wren's Nest Nature Reserve'den toplanmış Silüriyen'e tarihlenen deniz yatağı fosilleri -
 Yaygın görülen bir Üst Silüriyen öripteridi olan Eurypterus
Yaygın görülen bir Üst Silüriyen öripteridi olan Eurypterus -
 Pterygotus yaygın bir dağılıma sahip dev bir öripterid idi (bu illüstrasyonu verilen tür Devoniyen'deki P. anglicus türüdür)
Pterygotus yaygın bir dağılıma sahip dev bir öripterid idi (bu illüstrasyonu verilen tür Devoniyen'deki P. anglicus türüdür) -
 Trilobitler, Silüriyen esnasında hâlâ yaygın ve çeşitlilerdi. ABD merkezinde Calymene celebra fosilleri oldukça yaygın görülür.
Trilobitler, Silüriyen esnasında hâlâ yaygın ve çeşitlilerdi. ABD merkezinde Calymene celebra fosilleri oldukça yaygın görülür. -
 Halysites, bir tabulata mercanı olup Paleozoyik boyunca yaşamış ve nesli tükenmiş bir gruptur
Halysites, bir tabulata mercanı olup Paleozoyik boyunca yaşamış ve nesli tükenmiş bir gruptur -
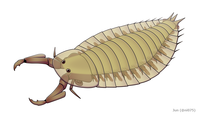 Parioscorpio Wisconsin'in Silüriyen zamanlarından kalma hakkında az şey bilinen bir eklem bacaklıdır.
Parioscorpio Wisconsin'in Silüriyen zamanlarından kalma hakkında az şey bilinen bir eklem bacaklıdır. -
 Dalmanites limulurus, New York'un Silüriyen katmanından bir örnektir
Dalmanites limulurus, New York'un Silüriyen katmanından bir örnektir -

-

-
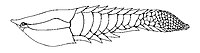 Kanada, Norveç ve ABD'nin Geç Silüriyen zamanlarından kalma bir zırhlı çenesiz balık cinsi olan Poraspis.
Kanada, Norveç ve ABD'nin Geç Silüriyen zamanlarından kalma bir zırhlı çenesiz balık cinsi olan Poraspis. -
 Entelognathus primordialis Geç Silüriyen'den kalma bir zırhlı balıktır.
Entelognathus primordialis Geç Silüriyen'den kalma bir zırhlı balıktır. -
 Qianodus[64] Çin'in Erken Silüriyen (Aroniyen) zamanlarından kalma diş bazlı kıkırdaklı balık cinsidir.
Qianodus[64] Çin'in Erken Silüriyen (Aroniyen) zamanlarından kalma diş bazlı kıkırdaklı balık cinsidir. -
![Shenacanthus[69] is jawed stem-chondrichthyan genus from the early Silurian (Telychian) of China](//upload.wikimedia.org/wikipedia/commons/thumb/6/6c/Shenacanthus_vermiformis.jpg/154px-Shenacanthus_vermiformis.jpg) Shenacanthus[65] Erken Silüriyen (Telisiyen) zamanlarından kalma çeneli, kök kıkırdaklı balık cinsidir.
Shenacanthus[65] Erken Silüriyen (Telisiyen) zamanlarından kalma çeneli, kök kıkırdaklı balık cinsidir. -
 Fanjingshania, Alt Silüriyen'den (Aroniyen) klimatid bir dikenli köpekbalığı olup parçalanmış deri iskeleti parçalarıyla tanınır
Fanjingshania, Alt Silüriyen'den (Aroniyen) klimatid bir dikenli köpekbalığı olup parçalanmış deri iskeleti parçalarıyla tanınır -











_1.jpg)




![Shenacanthus[69] is jawed stem-chondrichthyan genus from the early Silurian (Telychian) of China](/wiki/tr/Dosya:Shenacanthus_vermiformis.jpg)

{kind=link}
{kind=link}
Kaynakça
[değiştir | kaynağı değiştir]- ^ Jeppsson, L.; Calner, M. (2007). "The Silurian Mulde Event and a scenario for secundo—secundo events". Earth and Environmental Science Transactions of the Royal Society of Edinburgh. 93 (02): 135–154. doi:10.1017/S0263593300000377.
- ^ Munnecke, A.; Samtleben, C.; Bickert, T. (2003). "The Ireviken Event in the lower Silurian of Gotland, Sweden-relation to similar Palaeozoic and Proterozoic events". Palaeogeography, Palaeoclimatology, Palaeoecology. 195 (1): 99–124. doi:10.1016/S0031-0182(03)00304-3.
- ^ "Uluslararası Kronostratigrafik Çizelge" (PDF). www.stratigraphy.org. Uluslararası Stratigrafi Komisyonu.
- ^ Lucas, Sepncer (6 Kasım 2018). "The GSSP Method of Chronostratigraphy: A Critical Review". Frontiers in Earth Science. 6: 191. Bibcode:2018FrEaS...6..191L. doi:10.3389/feart.2018.00191
 .
. - ^ Holland, C. (June 1985). "Series and Stages of the Silurian System" (PDF). Episodes. 8 (2): 101-103. doi:10.18814/epiiugs/1985/v8i2/005 . 19 Ocak 2022 tarihinde kaynağından arşivlendi (PDF). Erişim tarihi: 11 Aralık 2020.
- ^ Chlupáč, Ivo; Hladil, Jindrich (January 2000). "The global stratotype section and point of the Silurian-Devonian boundary". CFS Courier Forschungsinstitut Senckenberg. Erişim tarihi: 7 Aralık 2020.
- ^ Haq, B. U.; Schutter, SR (2008). "A Chronology of Paleozoic Sea-Level Changes". Science. 322 (5898): 64-68. Bibcode:2008Sci...322...64H. doi:10.1126/science.1161648. PMID 18832639.
- ^ Bakınız:
- Murchison, Roderick Impey (1835). "On the Silurian system of rocks". Philosophical Magazine. 3rd series. 7 (37): 46-52. doi:10.1080/14786443508648654. 30 Ağustos 2023 tarihinde kaynağından arşivlendi. Erişim tarihi: 30 Ağustos 2023. From p. 48: " … I venture to suggest, that as the great mass of rocks in question, trending from south-west to north-east, traverses the kingdom of our ancestors the Silures, the term "Silurian system" should be adopted … "
- Wilmarth, Mary Grace (1925). Bulletin 769: The Geologic Time Classification of the United States Geological Survey Compared With Other Classifications, accompanied by the original definitions of era, period and epoch terms. Washington, D.C., U.S.: U.S. Government Printing Office. s. 80.
- ^ Sedgwick; Murchison, R.I. (1835). "On the Silurian and Cambrian systems, exhibiting the order in which the older sedimentary strata succeed each other in England and Wales". Report of the Fifth Meeting of the British Association for the Advancement of Science. § Notices and Abstracts of Miscellaneous Communications to the Sections. 5: 59-61. 30 Ağustos 2023 tarihinde kaynağından arşivlendi. Erişim tarihi: 30 Ağustos 2023.
- ^ Lapworth, Charles (1879). "On the tripartite classification of the Lower Palaeozoic rocks". Geological Magazine. 2nd series. 6 (1): 1-15. Bibcode:1879GeoM....6....1L. doi:10.1017/s0016756800156560. From pp. 13–14: "North Wales itself – at all events the whole of the great Bala district where Sedgwick first worked out the physical succession among the rocks of the intermediate or so-called Upper Cambrian or Lower Silurian system; and in all probability much of the Shelve and the Caradoc area, whence Murchison first published its distinctive fossils – lay within the territory of the Ordovices; … Here, then, have we the hint for the appropriate title for the central system of the Lower Palaeozoics. It should be called the Ordovician System, after this old British tribe."
- ^ The Gotlandian system was proposed in 1893 by the French geologist Albert Auguste Cochon de Lapparent (1839–1908): Lapparent, A. de (1893). Traité de Géologie (Fransızca). 2 (3. bas.). Paris, France: F. Savy. s. 748. 30 Ağustos 2023 tarihinde kaynağından arşivlendi. Erişim tarihi: 30 Ağustos 2023. From p. 748: "D'accord avec ces divisions, on distingue communément dans le silurien trois étages: l'étage inférieur ou cambrien (1) ; l'étage moyen ou ordovicien (2) ; l'étage supérieur ou gothlandien (3)." (In agreement with these divisions, one generally distinguishes, within the Silurian, three stages: the lower stage or Cambrian [1]; the middle stage or Ordovician [2]; the upper stage or Gotlandian [3].)
- ^ Barrande, Joachim (1852). Systême silurien du centre de la Bohême (Fransızca). Paris, France and Prague, (Czech Republic): (Self-published). ss. ix-x. 30 Ağustos 2023 tarihinde kaynağından arşivlendi. Erişim tarihi: 30 Ağustos 2023.
- ^ Forbes, Edward (1854). "Anniversary Address of the President". Quarterly Journal of the Geological Society of London. 10: xxii-lxxxi. 30 Ağustos 2023 tarihinde kaynağından arşivlendi. Erişim tarihi: 30 Ağustos 2023. See p. xxxiv.
- ^ a b c d e Munnecke, Axel; Calner, Mikael; Harper, David A.T.; Servais, Thomas (2010). "Ordovician and Silurian sea–water chemistry, sea level, and climate: A synopsis". Palaeogeography, Palaeoclimatology, Palaeoecology. 296 (3–4): 389-413. Bibcode:2010PPP...296..389M. doi:10.1016/j.palaeo.2010.08.001.
- ^ Yan, Guanzhou; Lehnert, Oliver; Männik, Peep; Calner, Mikael; Luan, Xiaocong; Gong, Fanyi; Li, Lixia; Wei, Xin; Wang, Guangxu; Zhan, Renbin; Wu, Rongchang (15 Kasım 2022). "The record of early Silurian climate changes from South China and Baltica based on integrated conodont biostratigraphy and isotope chemostratigraphy". Palaeogeography, Palaeoclimatology, Palaeoecology. 606: 111245. Bibcode:2022PPP...606k1245Y. doi:10.1016/j.palaeo.2022.111245. 7 Aralık 2022 tarihinde kaynağından arşivlendi. Erişim tarihi: 8 Aralık 2022.
- ^ a b Gambacorta, G.; Menichetti, E.; Trincianti, E.; Torricelli, S. (March 2019). "The Silurian climatic transition recorded in the epicontinental Baltica Sea". Palaeogeography, Palaeoclimatology, Palaeoecology. 517: 16-29. Bibcode:2019PPP...517...16G. doi:10.1016/j.palaeo.2018.12.016.
- ^ Young, Set A.; Benayoun, Emily; Kozik, Nevin P.; Hints, Olle; Martma, Tõnu; Bergström, Stig M.; Owens, Jeremy D. (15 Eylül 2020). "Marine redox variability from Baltica during extinction events in the latest Ordovician–early Silurian". Palaeogeography, Palaeoclimatology, Palaeoecology. 554: 109792. Bibcode:2020PPP...554j9792Y. doi:10.1016/j.palaeo.2020.109792.
- ^ Sproson, Adam D. (15 Şubat 2020). "Pacing of the latest Ordovician and Silurian carbon cycle by a ~4.5 Myr orbital cycle". Palaeogeography, Palaeoclimatology, Palaeoecology. 540: 109543. Bibcode:2020PPP...540j9543S. doi:10.1016/j.palaeo.2019.109543. Erişim tarihi: 8 Aralık 2022.
- ^ Trotter, Julie A.; Williams, Ian S.; Barnes, Christopher R.; Männik, Peep; Simpson, Andrew (February 2016). "New conodont δ18O records of Silurian climate change: Implications for environmental and biological events". Palaeogeography, Palaeoclimatology, Palaeoecology. 443: 34-48. Bibcode:2016PPP...443...34T. doi:10.1016/j.palaeo.2015.11.011.
- ^ Chaloner, William G. (2003). "The role of carbon dioxide in plant evolution". Evolution on Planet Earth: 65-83. doi:10.1016/B978-012598655-7/50032-X. ISBN 9780125986557.
- ^ Nealon, T.; Williams, D. Michael (30 Nisan 2007). "Storm-influenced shelf deposits from the silurian of Western Ireland: A reinterpretation of deep basin sediments". Geological Journal. 23 (4): 311-320. doi:10.1002/gj.3350230403.
- ^ Rasmussen, Christian M. Ø.; Kröger, Björn; Nielsen, Morten L.; Colmenar, Jorge (9 Nisan 2019). "Cascading trend of Early Paleozoic marine radiations paused by Late Ordovician extinctions". Proceedings of the National Academy of Sciences of the United States of America. 116 (15): 7207-7213. Bibcode:2019PNAS..116.7207R. doi:10.1073/pnas.1821123116. PMC 6462056 $2. PMID 30910963.
- ^ Capel, Elliot; Cleal, Christopher J.; Xue, Jinzhuang; Monnet, Claude; Servais, Thomas; Cascales-Miñana, Borja (August 2022). "The Silurian–Devonian terrestrial revolution: Diversity patterns and sampling bias of the vascular plant macrofossil record". Earth-Science Reviews. 231: 104085. Bibcode:2022ESRv..23104085C. doi:10.1016/j.earscirev.2022.104085.
- ^ Xue, Jinzhuang; Huang, Pu; Wang, Deming; Xiong, Conghui; Liu, Le; Basinger, James F. (May 2018). "Silurian-Devonian terrestrial revolution in South China: Taxonomy, diversity, and character evolution of vascular plants in a paleogeographically isolated, low-latitude region". Earth-Science Reviews. 180: 92-125. Bibcode:2018ESRv..180...92X. doi:10.1016/j.earscirev.2018.03.004. 9 Kasım 2022 tarihinde kaynağından arşivlendi. Erişim tarihi: 8 Aralık 2022.
- ^ Retallack, Gregory J. (June 2022). "Ordovician-Devonian lichen canopies before evolution of woody trees". Gondwana Research. 106: 211-223. Bibcode:2022GondR.106..211R. doi:10.1016/j.gr.2022.01.010. 15 Ocak 2023 tarihinde kaynağından arşivlendi. Erişim tarihi: 23 Kasım 2022.
- ^ Rittner, Don (2009). Encyclopedia of Biology. Infobase Publishing. s. 338. ISBN 9781438109992.
- ^ Rickards, R. B. (1 Mart 2000). "The age of the earliest club mosses: the Silurian Baragwanathia flora in Victoria, Australia". Geological Magazine. 137 (2): 207-209. Bibcode:2000GeoM..137..207R. doi:10.1017/S0016756800003800. 12 Kasım 2022 tarihinde kaynağından arşivlendi. Erişim tarihi: 8 Aralık 2022.
- ^ Lang, W.H.; Cookson, I.C. (1935). "On a flora, including vascular land plants, associated with Monograptus, in rocks of Silurian age, from Victoria, Australia". Philosophical Transactions of the Royal Society of London B. 224 (517): 421-449. Bibcode:1935RSPTB.224..421L. doi:10.1098/rstb.1935.0004.
- ^ Hueber, F.M. (1983). "A new species of Baragwanathia from the Sextant Formation (Emsian) Northern Ontario, Canada". Botanical Journal of the Linnean Society. 86 (1–2): 57-79. doi:10.1111/j.1095-8339.1983.tb00717.x.
- ^ Bora, Lily (2010). Principles of Paleobotany. Mittal Publications. ss. 36-37.
- ^ a b Niklas, Karl J. (1976). "Chemical Examinations of Some Non-Vascular Paleozoic Plants". Brittonia. 28 (1): 113-137. doi:10.2307/2805564. JSTOR 2805564.
- ^ Edwards, D.; Wellman, C. (2001), "Embryophytes on Land: The Ordovician to Lochkovian (Lower Devonian) Record", Gensel, P.; Edwards, D. (Ed.), Plants Invade the Land : Evolutionary and Environmental Perspectives, New York: Columbia University Press, ss. 3-28, ISBN 978-0-231-11161-4, p. 4
- ^ a b c Brookfield, M. E.; Catlos, E. J.; Suarez, S. E. (3 Ekim 2021). "Myriapod divergence times differ between molecular clock and fossil evidence: U/Pb zircon ages of the earliest fossil millipede-bearing sediments and their significance". Historical Biology (İngilizce). 33 (10): 2014-2018. doi:10.1080/08912963.2020.1762593. ISSN 0891-2963. 23 Haziran 2023 tarihinde kaynağından arşivlendi. Erişim tarihi: 30 Ağustos 2023.
- ^ Dunlop, Jason A. (1996). "A trigonotarbid arachnid from the Upper Silurian of Shropshire". Palaeontology. 39: 605-614. ISSN 0031-0239. 27 Haziran 2023 tarihinde kaynağından arşivlendi. Erişim tarihi: 30 Ağustos 2023.
- ^ Shear, William A.; Jeram, Andrew J.; Selden, Paul (1998). Centiped legs (Arthropoda, Chilopoda, Scutigeromorpha) from the Silurian and Devonian of Britain and the Devonian of North America. New York, NY: American Museum of Natural History. 27 Haziran 2023 tarihinde kaynağından arşivlendi. Erişim tarihi: 30 Ağustos 2023.
- ^ Jeram, Andrew J.; Selden, Paul A.; Edwards, Dianne (1990). "Land Animals in the Silurian: Arachnids and Myriapods from Shropshire, England". Science. 250 (4981): 658-61. Bibcode:1990Sci...250..658J. doi:10.1126/science.250.4981.658. PMID 17810866.
- ^ DiMichele, William A; Hook, Robert W (1992). "The Silurian". Behrensmeyer, Anna K. (Ed.). Terrestrial Ecosystems Through Time: Evolutionary Paleoecology of Terrestrial Plants and Animals. University of Chicago Press. ss. 207-10. ISBN 978-0-226-04155-1.
- ^ Paul Selden & Helen Read (2008). "The oldest land animals: Silurian millipedes from Scotland" (PDF). Bulletin of the British Myriapod & Isopod Group. 23: 36-37. 5 Temmuz 2016 tarihinde kaynağından arşivlendi (PDF). Erişim tarihi: 30 Ağustos 2023.
- ^ Wilson, Heather M.; Anderson, Lyall I. (2004). "Morphology and taxonomy of Paleozoic millipedes (Diplopoda: Chilognatha: Archipolypoda) from Scotland". Journal of Paleontology (İngilizce). 78 (1): 169-184. doi:10.1666/0022-3360(2004)078<0169:MATOPM>2.0.CO;2. ISSN 0022-3360. 27 Haziran 2023 tarihinde kaynağından arşivlendi. Erişim tarihi: 30 Ağustos 2023.
- ^ Suarez, Stephanie E.; Brookfield, Michael E.; Catlos, Elizabeth J.; Stöckli, Daniel F. (28 Haziran 2017). "A U-Pb zircon age constraint on the oldest-recorded air-breathing land animal". PLOS ONE. 12 (6): e0179262. doi:10.1371/journal.pone.0179262. ISSN 1932-6203. PMC 5489152 $2. PMID 28658320.
- ^ Rong, Jia-yu; Shen, Shu-zhong (1 Aralık 2002). "Comparative analysis of the end-Permian and end-Ordovician brachiopod mass extinctions and survivals in South China". Palaeogeography, Palaeoclimatology, Palaeoecology. 188 (1–2): 25-38. Bibcode:2002PPP...188...25R. doi:10.1016/S0031-0182(02)00507-2. 16 Nisan 2023 tarihinde kaynağından arşivlendi. Erişim tarihi: 16 Nisan 2023.
- ^ Rong, Jiayu; Zhan, Renbin (October 1999). "Chief sources of brachiopod recovery from the end Ordovician mass extinction with special references to progenitors". Science in China Series D: Earth Sciences. 42 (5): 553-560. Bibcode:1999ScChD..42..553R. doi:10.1007/BF02875250. 16 Nisan 2023 tarihinde kaynağından arşivlendi. Erişim tarihi: 16 Nisan 2023.
- ^ Huang, Bing; Rong, Jiayu; Cocks, L. Robin M. (1 Şubat 2012). "Global palaeobiogeographical patterns in brachiopods from survival to recovery after the end-Ordovician mass extinction". Palaeogeography, Palaeoclimatology, Palaeoecology. 317-318: 196-205. Bibcode:2012PPP...317..196H. doi:10.1016/j.palaeo.2012.01.009. 16 Nisan 2023 tarihinde kaynağından arşivlendi. Erişim tarihi: 16 Nisan 2023.
- ^ a b Huang, Bing; Jin, Jisuo; Rong, Jia-Yu (15 Mart 2018). "Post-extinction diversification patterns of brachiopods in the early–middle Llandovery, Silurian". Palaeogeography, Palaeoclimatology, Palaeoecology. 493: 11-19. Bibcode:2018PPP...493...11H. doi:10.1016/j.palaeo.2017.12.025. 23 Kasım 2022 tarihinde kaynağından arşivlendi. Erişim tarihi: 23 Kasım 2022.
- ^ Jia-Yu, Rong; Harper, David A. T. (31 Ocak 2000). "Brachiopod survival and recovery from the latest Ordovician mass extinctions in South China". Geological Journal. 34 (4): 321-348. doi:10.1002/(SICI)1099-1034(199911/12)34:4<321::AID-GJ809>3.0.CO;2-I. 21 Nisan 2023 tarihinde kaynağından arşivlendi. Erişim tarihi: 16 Nisan 2023.
- ^ Anstey, Robert L.; Pachut, Joseph F.; Tuckey, Michael E. (8 Nisan 2016). "Patterns of bryozoan endemism through the Ordovician-Silurian transition". Paleobiology. 29 (3): 305-328. doi:10.1666/0094-8373(2003)0292.0.CO;2. 16 Nisan 2023 tarihinde kaynağından arşivlendi. Erişim tarihi: 16 Nisan 2023.
- ^ Zapalski, Mikołaj K.; Vinn, Olev; Toom, Ursula; Ernst, Andrej; Wilson, Mark A. (16 Eylül 2022). "Bryozoan–cnidarian mutualism triggered a new strategy for greater resource exploitation as early as the Late Silurian". Scientific Reports. 12 (1): 15556. Bibcode:2022NatSR..1215556Z. doi:10.1038/s41598-022-19955-2. PMC 9481587 $2. PMID 36114227.
- ^ Claussen, Anna Lene; Munnecke, Axel; Ernst, Andrej (2 Mart 2021). "Bryozoan-rich stromatolites (bryostromatolites) from the Silurian of Gotland and their relation to climate-related perturbations of the global carbon cycle". Sedimentology. 69 (1): 162-198. doi:10.1111/sed.12863. 16 Nisan 2023 tarihinde kaynağından arşivlendi. Erişim tarihi: 16 Nisan 2023.
- ^ Liljedahl, Louis (January 1985). "Ecological aspects of a silicified bivalve fauna from the Silurian of Gotland". Lethaia. 18 (1): 53-66. doi:10.1111/j.1502-3931.1985.tb00684.x. 16 Nisan 2023 tarihinde kaynağından arşivlendi. Erişim tarihi: 16 Nisan 2023.
- ^ Claussen, Anna Lene; Munnecke, Axel; Wilson, Mark A.; Oswald, Irina (22 Nisan 2019). "The oldest deep-boring bivalves? Evidence from the Silurian of Gotland (Sweden)". Facies. 65 (3). doi:10.1007/s10347-019-0570-7. 16 Nisan 2023 tarihinde kaynağından arşivlendi. Erişim tarihi: 16 Nisan 2023.
- ^ Cherns, Lesley (2 Ocak 2007). "Early Palaeozoic diversification of chitons (Polyplacophora, Mollusca) based on new data from the Silurian of Gotland, Sweden". Lethaia. 37 (4): 445-456. doi:10.1080/00241160410002180. 30 Ağustos 2023 tarihinde kaynağından arşivlendi. Erişim tarihi: 16 Nisan 2023.
- ^ a b Vinn, Olev; Wilson, Mark A.; Madison, Anna; Kazantseva, Elizaveta; Toom, Ursula (26 Temmuz 2022). "First Symbiotic Association Between Hederelloids and Rugose Corals (Latest Silurian of Saaremaa, Estonia)". PALAIOS. 37 (7): 368-373. Bibcode:2022Palai..37..368V. doi:10.2110/palo.2022.005. 16 Mayıs 2023 tarihinde kaynağından arşivlendi. Erişim tarihi: 16 Nisan 2023.
- ^ Wittmer, Jacalyn M.; Miller, Arnold I. (1 Aralık 2011). "Dissecting the global diversity trajectory of an enigmatic group: The paleogeographic history of tentaculitoids". Palaeogeography, Palaeoclimatology, Palaeoecology. 312 (1–2): 54-65. Bibcode:2011PPP...312...54W. doi:10.1016/j.palaeo.2011.09.009. 16 Nisan 2023 tarihinde kaynağından arşivlendi. Erişim tarihi: 16 Nisan 2023.
- ^ Wei, Fan; Gong, Yiming; Yang, Hao (1 Kasım 2012). "Biogeography, ecology and extinction of Silurian and Devonian tentaculitoids". Palaeogeography, Palaeoclimatology, Palaeoecology. 358-360: 40-50. Bibcode:2012PPP...358...40W. doi:10.1016/j.palaeo.2012.07.019. 16 Nisan 2023 tarihinde kaynağından arşivlendi. Erişim tarihi: 16 Nisan 2023.
- ^ Wei, Xin; Zhan, Ren-Bin (March 2018). "A late Rhuddanian (early Llandovery, Silurian) trilobite association from South China and its implications". Palaeoworld. 27 (1): 42-52. doi:10.1016/j.palwor.2017.05.008. 16 Nisan 2023 tarihinde kaynağından arşivlendi. Erişim tarihi: 16 Nisan 2023.
- ^ Adrain, Jonathan M.; Westrop, Stephen R.; Chatterton, Brian D. E.; Ramsköld, Lars (8 Şubat 2016). "Silurian trilobite alpha diversity and the end-Ordovician mass extinction". Paleobiology. 26 (4): 625-646. doi:10.1666/0094-8373(2000)0262.0.CO;2. 16 Nisan 2023 tarihinde kaynağından arşivlendi. Erişim tarihi: 16 Nisan 2023.
- ^ Peters, Shanan E.; Ausich, William I. (8 Nisan 2016). "A sampling-adjusted macroevolutionary history for Ordovician-Early Silurian crinoids". Paleobiology. 34 (1): 104-116. doi:10.1666/07035.1. 16 Nisan 2023 tarihinde kaynağından arşivlendi. Erişim tarihi: 16 Nisan 2023.
- ^ Wright, David F.; Toom, Ursula (30 Ağustos 2017). "New crinoids from the Baltic region (Estonia): fossil tip-dating phylogenetics constrains the origin and Ordovician–Silurian diversification of the Flexibilia (Echinodermata)". Palaeontology. 60 (6): 893-910. doi:10.1111/pala.12324. 16 Nisan 2023 tarihinde kaynağından arşivlendi. Erişim tarihi: 16 Nisan 2023.
- ^ Ausich, William I.; Deline, Bradley (15 Kasım 2012). "Macroevolutionary transition in crinoids following the Late Ordovician extinction event (Ordovician to Early Silurian)". Palaeogeography, Palaeoclimatology, Palaeoecology. 361-362: 38-48. Bibcode:2012PPP...361...38A. doi:10.1016/j.palaeo.2012.07.022. 16 Nisan 2023 tarihinde kaynağından arşivlendi. Erişim tarihi: 16 Nisan 2023.
- ^ Zong, Rui-wen; Gong, Yi-ming (1 Kasım 2020). "Discovery of scyphocrinoid loboliths in western Junggar, Xinjiang, NW China: Implications for scyphocrinoid paleobiogeography and identification of the Silurian–Devonian boundary". Palaeogeography, Palaeoclimatology, Palaeoecology. 557: 109914. Bibcode:2020PPP...557j9914Z. doi:10.1016/j.palaeo.2020.109914. 7 Aralık 2022 tarihinde kaynağından arşivlendi. Erişim tarihi: 16 Nisan 2023.
- ^ Vinn, O.; wilson, M.A.; Mõtus, M.-A. (2014). "Symbiotic endobiont biofacies in the Silurian of Baltica". Palaeogeography, Palaeoclimatology, Palaeoecology. 404: 24-29. Bibcode:2014PPP...404...24V. doi:10.1016/j.palaeo.2014.03.041. 11 Aralık 2019 tarihinde kaynağından arşivlendi. Erişim tarihi: 11 Haziran 2014.
- ^ Vinn, O.; Mõtus, M.-A. (2008). "The earliest endosymbiotic mineralized tubeworms from the Silurian of Podolia, Ukraine". Journal of Paleontology. 82 (2): 409-414. Bibcode:2008JPal...82..409V. doi:10.1666/07-056.1. 8 Mayıs 2020 tarihinde kaynağından arşivlendi. Erişim tarihi: 11 Haziran 2014.
- ^ Zatoń, Michał; Wrzołek, Tomasz (15 Ekim 2020). "Colonization of rugose corals by diverse epibionts: dominance and syn vivo encrustation in a Middle Devonian (Givetian) soft-bottom habitat of the Holy Cross Mountains, Poland". Palaeogeography, Palaeoclimatology, Palaeoecology. 556: 109899. Bibcode:2020PPP...556j9899Z. doi:10.1016/j.palaeo.2020.109899. 16 Nisan 2023 tarihinde kaynağından arşivlendi. Erişim tarihi: 16 Nisan 2023.
- ^ Andreev, Plamen S.; Sansom, Ivan J.; Li, Qiang; Zhao, Wenjin; Wang, Jianhua; Wang, Chun-Chieh; Peng, Lijian; Jia, Liantao; Qiao, Tuo; Zhu, Min (September 2022). "The oldest gnathostome teeth". Nature (İngilizce). 609 (7929): 964–968. Bibcode:2022Natur.609..964A. doi:10.1038/s41586-022-05166-2. ISSN 1476-4687. PMID 36171375. 21 Ekim 2022 tarihinde kaynağından arşivlendi. Erişim tarihi: 30 Ağustos 2023.
- ^ Zhu, You-an; Li, Qiang; Lu, Jing; Chen, Yang; Wang, Jianhua; Gai, Zhikun; Zhao, Wenjin; Wei, Guangbiao; Yu, Yilun; Ahlberg, Per E.; Zhu, Min (2022). "The oldest complete jawed vertebrates from the early Silurian of China". Nature (İngilizce). 609 (7929): 954–958. doi:10.1038/s41586-022-05136-8. ISSN 1476-4687. PMID 36171378. 19 Ekim 2022 tarihinde kaynağından arşivlendi. Erişim tarihi: 30 Ağustos 2023.
Genel
- Emiliani, Cesare. (1992). Planet Earth : Cosmology, Geology, & the Evolution of Life & the Environment. Cambridge University Press. (Paperback Edition 0-521-40949-7)
- Mikulic, DG, DEG Briggs, and J Kluessendorf. 1985. A new exceptionally preserved biota from the Lower Silurian of Wisconsin, USA. Philosophical Transactions of the Royal Society of London, 311B:75-86.
- Moore, RA; Briggs, DEG; Braddy, SJ; Anderson, LI; Mikulic, DG; Kluessendorf, J (2005). "A new synziphosurine (Chelicerata: Xiphosura) from the Late Llandovery (Silurian) Waukesha Lagerstatte, Wisconsin, USA". Journal of Paleontology. 79 (2): 242-250. doi:10.1666/0022-3360(2005)079<0242:anscxf>2.0.co;2.
Dış bağlantılar
[değiştir | kaynağı değiştir]- Silüriyen - Biltek Tübitak
- Ogg, Jim (June 2004), Overview of Global Boundary Stratotype Sections and Points (GSSP's)
- Palaeos: Silüriyen
- UCMP Berkeley: 17 Haziran 2010 tarihinde Wayback Machine sitesinde arşivlendi. Silüriyen
- Paleoportal: Eyaletten eyalete ABD'deki Silüriyen katmanları 15 Mart 2021 tarihinde Wayback Machine sitesinde arşivlendi.
- USGS: Silüriyen ve Devoniyen Kayaçları (ABD)
- "International Commission on Stratigraphy (ICS)". Geologic Time Scale 2004. 20 Eylül 2005 tarihinde kaynağından arşivlendi. Erişim tarihi: 19 Eylül 2005.
- Silüriyen fosillerine örnekler 12 Ocak 2010 tarihinde Wayback Machine sitesinde arşivlendi.
- Silüriyen - GeoWhen Veritabanı 30 Ağustos 2023 tarihinde Wayback Machine sitesinde arşivlendi.
- Silüriyen (Kronostratigrafi cetveli) 11 Mayıs 2022 tarihinde Wayback Machine sitesinde arşivlendi.